
|
Anterior to the midbrain the interbrain lies between the cerebral hemispheres. The interbrain includes the optic thalamus, the corpora mammillaria, corpora geniculata, pineal gland, and most of the third ventricle. Lateral to the thalamus lie the corpora striata. These bodies are properly a part of the cerebral hemispheres, but for the convenience of discussion they are included in this group. The
ganglia grouped around the base of the brain have been studied for a long
time, yet their physiological significance is as poorly understood as is
any part of the nervous system. Part of the difficulty in study lies
in the fact that it is very difficult to reach them for purposes of experiment
without disturbing the cortex or the fiber tracts which relate the cortex
to other parts of the nervous system. The location of these ganglia
between many tracts, associating the areas of the cortex with one another
and with the lower centers, accounts for the fat that so few clinic cases
are recorded in which uncomplicated lesions of the corpora striata or the
optic thalamus or others of these centers have been found. What experimental
and clinical evidence is at hand presents many conflicting and apparently
irreconcilable factors. Further study is needed. It is especially
desirable that pathological brains should be studied by recent methods
of neurological technique, and that such findings should be considered
in the light of the symptoms observed before death.
The optic thalamus includes a number of centers of varying functions and relationships. Nissl described twenty different nuclei in the thalamus of the rabbit. Later authors do not verify all of these. With our present incomplete knowledge of these centers it is wiser to follow their example and discuss the centers in groups, leaving their further differentiation for later investigations. The thalamus is an ovoid body lying medial from the striate body and lateral and inferior to the third ventricle. It forms part of the lateral wall of this ventricle. The word thalamus means bed, and it is so called because it presents a couchlike prominence into the ventricle. At its posterior end the pulvinar forms a swelling, which is so called because it is supposed to resemble the pillow of the thalamus, or bed. Beneath the pulvinar are two other swellings, the lateral geniculate body and the median geniculate body. Anteriorly, upon the lateral area of the superior surface of the thalamus appears the anterior tubercle, beneath which lies the anterior nucleus of the thalamus. The thalamus itself is covered by white and gray matter. The gray matter is continuous with the gray matter lining the ventricles. The white matter is composed of the fibers entering and leaving the thalamus. These make up a sheath for the body, and prolongations from this sheath form various partitions, which are more or less complete, and which divide the inner gray matter into many different centers or nuclei. The gray matter of each thalamus is continuous with that of its fellow beneath the third ventricle. The
nuclei of the thalamus are divided into the following groups: The
lateral nucleus, medial nucleus, anterior nucleus, the nucleus of the pulvinar
and the nucleus habenulae.
This nucleus lies beneath the trigonum habenulae. It is of interest because of its relation to the rhinencephalon. It receives olfactory impulses by way of the thalamic striae. Fibers from the peduncles of the pineal gland also enter this nucleus. The
axons of the cells of the nucleus habenulae pass in part to the pineal
gland, but for the most part these fibers make up the fasciculus retroflexus
(Meynerts bundle), which passes downward through the tegmentum to the
ganglion interpedunculare. From this ganglion the fibers pass to
the nuclei of the reticular region, and thence to the motor nuclei of the
cranial and spinal nerves. The reflex actions associated with the
olfactory impulses are thus, in part, controlled.
This must not be confused with the anterior nucleus of older authors. It lies on the anterior portion of the dorsal aspect of the thalamus. It is completely inclosed by white fibers, which are continuous with the stratum zonale of the thalamus. It gives fibers to the corpus albicans by way of the fasciculus thalamo-mammillaris (bundle of Vicq dAzyr), and receives from it the axons of the cells of the corpora mammillaria (corpora albicantia). Since these bodies receive the fornix fibers, the anterior tubercle of the thalamus brings into functional relationship the thalamus and the hippocampal region. The connections thus secured are functional in the coordination of the olfactory impulses. The pulvinar is a mass of gray matter continuous with the lateral nucleus of the thalamus. It receives about one-fifth of the optic tract fibers. The axons of the cells of the pulvinar pass for the most part by way of the optic radiations to the cortex of the occipital lobe, but others pass also to the superior quadrigeminate by way of the superior brachium. Others of its fibers pass centripetally in the optic tracts to the retina. Injury of the puplvinar causes homonymous hemianopsia. This is the largest nucleus of the thalamus. It is divided into a number of smaller nuclei, which may or may not be related in function. The lateral nucleus extends from the anterior portion of the thalamus to the posterior, and should be used to include the nucleus of Luys (thalamic) or the central median nucleus, and the arcuate nucleus. The lateral nucleus fuses with the pulvinar at its posterior region. The lateral nucleus receives fibers from the following sources: I. The median fillet terminates in this nucleus. A few of the fillet fibers pass without relay into the optic radiations. The impulses concerned in the appreciation of touch, muscular effort and the common sensations are thus transmitted to the sensori-motor areas of the cortex. II. The spino-thalamic tract carries to the thalamus the impulses of temperature changes, pain and touch. Part of these fibers also may enter the radiations without relay. III. Fibers from the medial longitudinal bundle enter the lateral nucleus. These fibers are homologous with the fibers from the posterior horn cells and the cells of the central gray matter of the cord. They transmit to the thalamus the impulses of pain and temperature, and perhaps also of touch. IV. The fibers of the brachium conjunctivum which failed to stop in the neighborhood of the red nucleus terminate in the thalamus. This represents a step in the indirect sensory conduction path. The viscero-sensory impulses, and those of touch, pain, temperature and muscular effort, are thus transmitted in part. Axons of cells of the red nucleus join the brachium fibers and in part terminate in the lateral nucleus. V. Descending fibers from the cortex enter the lateral nucleus with the radiations. The functions of these descending fibers are not known. VI. Fibers from the putamen and the caudate nucleus enter the thalamus, and either immediately or after synapsis with cells of the median or other nuclei of the thalamus, carry impulses to the lateral nucleus. The lateral nucleus is thus an important relay station in the sensory pathway to the cortex. Its fibers are given off as follows: I. The chief destination of the fibers is to the cortex of the postcentral convolutions. II. Fibers pass to the corpora striatum, and to other not very well-studied terminations in the related ganglia of the midbrain, the interbrain, the subthalamic and the hypothalamic regions. Injury
of the optic thalamus of one side causes anesthesia, analgesia and ataxia
of the opposite side of the body, chiefly, but to a certain extent on the
same side of the body.
The median nucleus of the thalamus lies near the median line. It is continuous by gray matter with its fellow of the opposite side, but it is separated from the other thalamic nuclei by a sheet of white fibers. It is continuous also with the hypothalamic gray matter. The median nucleus receives fibers as follows: I. The strio-thalamic tract seems to send certain of its fibers, if not its chief bundle, to this nucleus. II. Descending fibers from the operculum and the frontal lobes terminate in this nucleus. III. Association fibers from other thalamic nuclei and probably from neighboring gray matter terminate in this nucleus. The median nucleus seems to be a relay station in the descending as well as in the ascending pathway. The axons of its cells pass to the following destinations, probably: I. To the cortex of the frontal lobes and the operculum. II. To other thalamic and neighboring centers. III.
To the descending thalamo-spinal tract. This tract seems to pass
with the rubro-spinal tract through the length of the cord, terminating
at all levels in the region of the central part of the crescent of gray
matter. Probably fibers are given to the motor nuclei of the cranial
nerves also.
The thalamus is one of the oldest of structures. A structure homologous with the thalamus of the mammal is found in the lowest of vertebrates The functions are variable in different classes of vertebrates, and this changing of function is doubtless due, as in other parts of the nervous system, to the parallel processes of cephalization and specialization. In the earlier fishes there is no exact division between the midbrain and the interbrain. The fillets, both medial and lateral, terminate in the undifferentiated tectum mesencephali. In the bony fishes, and in all vertebrates above these, as well as in certain of the lower forms, the thalamus can be distinguished as a separate structure. The nucleus habenulae presents the rare history of a nucleus present in a certain degree of development and with certain morphological relationships in the lower vertebrates, retaining its morphological and functional relationships through varying degrees of cephalization and specialization on the part of the rest of the nervous system, yet itself neither becoming more specialized, nor more complex, nor yet losing place or becoming in any wise reminiscent. From the beginning a center associated with olfactory impulses, and transmitting these to the motor centers, it is found in man performing the same functions in practically the same manner. It is true that its position in the scheme of the physiological relationships is apparently less important in the higher vertebrates than in the lower. Yet even in man there is reason to believe that the reflexes aroused by olfactory impulses are more profound and more efficient than has heretofore been believed. The nucleus of the anterior tubercle also has a history of comparatively uncephalized and unspecialized existence. It is, and has been, associated with the olfactory impulses and their coordination in modifying the movements of the body. The pushing forward of the visual associations is of interest in this connection. The termination of the larger part of the optic fibers in the region of the midbrain was followed by the development of collaterals from these fibers which entered the lateral geniculate body. The main fibers of the optic tract to the superior colliculus became less functional and less numerous, while the fibers, once collaterals to the lateral geniculate body, became more and more efficient. Later the same processes have resulted in the formation of yet another termination, in the pulvinar of the thalamus. The auditory neurons have like relationships. First the lateral fillet terminated in the roof of the midbrain, indifferently. Then the posterior colliculus claimed the greater share of the auditory fibers, and this became the chief center for the coordination of the auditory impulses in the control of the movements of the body. Later the development of the anterior centers resulted in the passing of numbers of fibers and collaterals from the auditory fillet to the median geniculate body. This body thus became the most important center in the pathway of the auditory impulses toward the cortex. The development of the lateral nucleus is yet unknown. In the lower vertebrates the median lemnicus terminates in the thalamus, but nothing is known of a thalamo-cortical tract. In monotremes there is found a well-marked tract from the thalamus to the cortex, carrying cutaneous sensory impulses. The median nucleus is one of the older nuclei. First being chiefly concerned in the transmission of impulses from the striate bodies to the lower motor centers, it has become one of the important centers for the coordination of complex reactions of a reflex or pseudo-reflex nature. It should be noted in the case of the thalamus, as in the case of the quadrigeminates and the cerebellum, that the same processes which lead to the function of any organ being susperseded by higher centers, lead also to the development of new coordinations, sometimes only distantly related to the older and more general functions. The
place of the thalamic centers in the emotional reactions is a matter of
dispute. The median nucleus seems to be concerned in these reactions.
Stimulation of the sub-thalamic region in the cat and the dog initiate
the movements characteristic of anger. It is not possible to localize
the exact center stimulated in such experiments.
The medial geniculate body lies upon the inferior aspect of the thalamus, medially to the lateral geniculate body. It appears to be the terminal ganglion of the central root of the optic tracts. It does receive the fibers of Fuddens commissure, which run with the optic tracts posterior to the chiasma, but which are not functionally related to the visual centers. The medial geniculate body contains small cells of spindle or fusiform outline, with few small dendrites It does not present a laminated appearance on section, and appears to be of simple structure. It receives fibers as follows: I. Axons of cells of the auditory nuclei, the superior olivary nuclei, and the nuclei of the trapezoid body reach this body by way of the lateral fillet. In this way the impulses concerned in consciousness of hearing are carried to the cortex. II. Axons of cells of the inferior quadrigeminates pass by way of the inferior brachium to this body. In this way also some of the auditory impulses are carried. III. Axons of the lateral geniculate body of the opposite side, and probably of the corpus striatum of the opposite side, by way of part of the lenticular loop fibers, pass by way of Guddens commissure to each lateral geniculate body. IV. Axons of cells in the centers of the thalamus probably end in the lateral geniculate body, bringing into coordinate function these centers. V. Axons of cells of the cortex of the auditory areas of the temporal lobes pass by way of the acustic radiation to the lateral geniculate body. Impulses carried by this path seem to be essential to the maintenance of normal nutritive conditions of the auditory structures, particularly the cochlea. The axons of the cells of the median genticulate body pass to the following structures: I. Axons pass in the acustic radiations to the cerebral cortex of the first and second temporal convolutions, the so-called auditory area. Thus the impulses concerned in conscious hearing are carried. II. Axons pass to the inferior quadrigeminates by way of the inferior brachium. The descending cortical impulses are thus transmitted peripherally in part. III. Axons descend along the path of the lateral fillet. In this way also the descending impulses are transmitted peripherally. IV. Axons pass by way of Guddens commissure to the opposite median geniculate body, and perhaps to the striate body of the opposite side. V. Axons pass to the thalamic nuclei of related function. VI. Axons pass to the red nucleus and related ganglia. The medial geniculate bodies are not subject to disease, but are affected by abnormal processes of neighboring structures. Injury of either median geniculate body produces deafness of both earschiefly of the opposite side, according to some authors, but equally, according to others. Usually the injury of either is associated with injury of the other. Stimulation
of the median geniculate bodies of the cat causes movements of the eyes
and the ears, provided the inferior connections are not injured.
The presence or absence of the cortex seems to have no effect upon the
results of stimulation of these bodies under the conditions of our experiments.
This is a swelling or protuberance upon the inferior and lateral aspect of the thalamus. It is phylogenetically ancient, being found in the lower fishes, but it seems of very little importance below mammals. This nucleus contains large, multipolar, pigmented nerve cells, whose dendrites branch freely. The nerve fibers which enter it pass through its substance in sheets or layers, and thus give the nucleus a striated appearance something like that of the anterior quadrigeminate body. The lateral geniculate body receives fibers form the following sources: I. About eighty per cent of the optic-tract fibers terminate in this nucleus and the pulvinar. In man, about one-half of these fibers are from the homo-lateral retina, and about half from the contra-lateral retina. In lower vertebrates the number of crossed fibers is much greater; in animals whose eyes are situated very far at the side, so that the two fields of vision do not overlap, the lateral geniculate body receives only contra-lateral retinal fibers. II. Fibers from the pulvinar and perhaps other thalamic centers bring the activity of the nuclei associated with sight sensations into coordinate function. III. Descending fibers from the occipital cortex terminate among the cells of the geniculate body. The impulses thus derived seem in some way to be essential to the nutrition of the ocular structures, and especially of the retina, but the manner of their activity is not known. IV. Axons pass to the red nucleus and related ganglia. The lateral geniculate body is not itself very subject to disease, but injury of neighboring parts of the brain often affect it. When it is injured, bilateral homonymous hemianopsia is produced. Experimental
stimulation of the body after ablation of the midbrain produces no effects.
With an intact midbrain stimulation of the lateral geniculate of the cat
causes movements of both the eyes and the ears of both sides. Ablation
of the cortex does not seem to affect the movements resulting from this
stimulation.
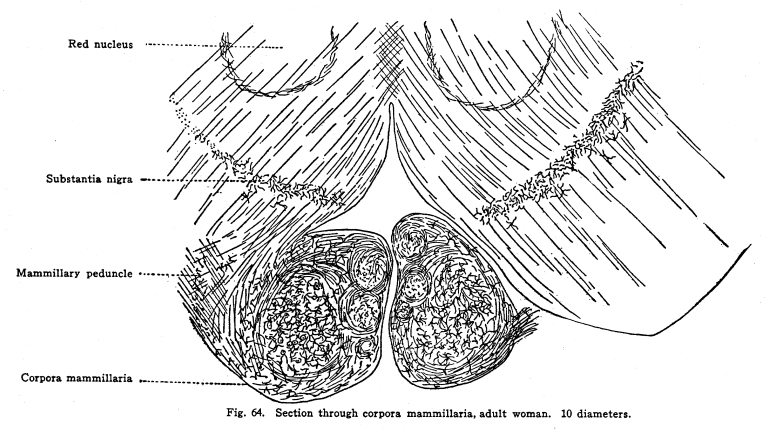
These bodies lie in front of the posterior perforated space and behind the optic commissure. Each corpus is the size of half a small pea, and about the same shape. The structure of this body is rather more complex than at first appears. (Fig. 64.) Its outer layer is composed of white matter, the descending fornix fibers. These pass around the lateral surface of the body, beneath it, and upward on its median aspect. As the fibers thus encircle the body they pass into its substance, a few at a time, until at the medial upper part of the body all the encircling fibers have entered the inner gray matter. The entering fibers, after plunging into the gray matter, form a series of whirls, which inclose within them a number of fairly large multipolar cells with long but not very freely branching dendrites. The axons of these cells pass with the entering fibers into the deeper gray matter of the corpus. The central portion of the median nucleus is composed of gray matter, together with the fibers terminating and originating therein. The cells of this nucleus are rather smaller than those just mentioned, their dendrites are shorter, and they branch more freely. The entering fibers break up into a number of fibrillae, which form large baskets around the intrinsic cells. Each fiber may enter into the formation of several baskets, and each basket may receive fibrillae from several fibers (I have found four fibrillae entering one basket, and one fiber giving fibrillae to two baskets, in several instances.) (Fig. 65.) Fig. 64. Section through corpora mammillaria, adult woman. 10 diameters. Red nucleus; Substantia nigra; Mammillary peduncle; Corpora mammillaria. Fig. 65. Calls from corpora mammillaria. 800 diameters. These baskets are much larger than the bodies of the cells which they inclose, and are composed of an extremely fine network, so that in sections one often finds part of the network with no cell body apparent within it. This gives something the appearance of the olfactory glomeruli to the sections. These baskets are not arranged in any apparent order, as are the olfactory glomeruli, but lie scattered all over the central portion of the median nucleus. The axons of the cells of the median nucleus form the principal mammillary fasciculus. The fibers of this bundle pass upward and bifurcate. One branch enters the anterior nucleus of the thalamus. This is the thalamo-mammillary bundle (bundle of Vicq dAzyr). Probably some fibers, axons of the thalamic cells, pass downward with this bundle into the corpora. The other branch, the pedunculo-mammillary bundle, passes backward toward the tegmentum. It runs near the median longitudinal bundle through the tegmentum, and gives collaterals or axons to the substantia nigra and the red nucleus. Both branches of the principal mammillary bundle are concerned in carrying impulses which relate the bodily movements in answer to the olfactory impulses. The
lateral nucleus of the corpora mammillaria is smaller. It receives
a part of the descending fornix fibers. These form synapses with
multipolar cells of median size, with rather long but not very freely branching
dendrites. The axons of some of these cells pass with the remaining
fornix fibers to the median nucleus. Other axons enter the pedunculo-mammillary
bundle as its basilar portion, and pass with it through the tegmentum.
These fibers seem to terminate in the central gray matter of the midbrain,
and through this are brought into relationship with the viscero-motor nulei
of the medulla. Perhaps the somatic cranial nerve nuclei are also
under the influence of the impulses carried by this tract, and indirectly
the centers of the cord may also be affected thereby.
The red nucleus, or nucleus rubber (nucleus of Stilling), lies beneath the aqueduct and the third ventricle. It receives its name from the fact that it is unusually freely supplied with blood, so that in the fresh specimen it presents a distinctly reddish appearance. This nucleus contains extremely large cells, whose dendrites attain great length and branch very freely. These cells are inclosed by the branching incoming axons and collaterals, and by their own dendritic and collateral branchings, as well as by the axons of the Golgi cells of Type II. The red nucleus receives fibers from the following sources: I. From the nucleus dentatum by way of the brachium conjunctivum. II. From the lateral and the medial fillet. III. Descending fibers from the cerebral cortex, chiefly of the frontal, parietal and occipital regions. IV. Association fibers from the neighboring gray matter, the striate bodies and the thalamus, especially the median nucleus of the thalamus. The axons of the cells of the red nucleus pass as follows: I. To the nucleus dentatus by way of the brachium conjunctivum. II. To the thalamus, and to the cerebral cortex of the somesthetic area. III. To the neighboring gray matter. IV. To the motor nuclei of the cranial nerves, and to the centers both visceral and somatic in the pons, medulla and midbrain, and to the center of the gray crescent of the spinal cord of all levels, by way of the rubro-spinal tract. The
red nucleus must be considered as one of the centers for the control of
the instinctive and emotional reactions.
The tuber cinereum lies immediately posterior to the optic chiasma. Its gray surface is directly continuous with the posterior perforated space of the interpeduncular region. Its protrusion beyond the rest of the gray matter is due to the outgrowth of the infundiculum. The infundibulum is a tube, an outgrowth from the inferior aspect of the third ventricle. It passes obliquely downward into the hypophesis cerebri, or pituitary body. The pituitary body, or hypophysis cerebri, is found in all vertebrates, but very much larger in the lower types. The posterior lobe has been studied thoroughly by Berkeley. He describes three chief varieties of cells, large and small pyramidal, very much like those of other parts of the nervous system, and smaller cells with freely-branching dendrites, and three or four small, slender axons, which terminate in forming synapses with other cells of the same structures. All three classes of cells send axons toward the infundibulum, but neither Berkeley nor any one else has described these axons as terminating in other parts of the nervous system. Gland cells are also found in the posterior lobe. The anterior lobe of the pituitary body is not of nervous origin. It is developed as an outgrowth from the pharyngeal region, comes into anatomical relationship with the posterior lobe, and remains in that place. Both lobes are very freely supplied with blood vessels, which are plentifully innervated by sympathetic fibers. The glandular tissues of the anterior lobe are supplied with sympathetic fibers, which have a termination which suggests that they are secretory in function. The pituitary body may be diseased, but not frequently. It may be hypertrophied, atrophied, or subject to tumors. When it is diseased the patient may suffer from no symptoms referable to the gland, or he may suffer a peculiar overgrowth of certain bones, as in the disease called acromegalia. Experimental
stimulation of the gland gives no effects. Extracts from it may affect
blood pressure. The presence of colloid material in the acini suggests
a relationship with the thyroid gland.
The corpus striatum is so called from its being penetrated by layers of the white fibers, which give its gray matter a striated appearance. It is a body of about the shape and size of a hens egg. It measures about two and a half inches in its antero-posterior diameter, and an inch and a half or an inch and a quarter in its other diameters. It
is divided into two divisions by the internal capsule, the caudate nucleus
and the lenticular nucleus.
The caudate nucleus forms a part of the floor of the lateral part of the lateral ventricle. Its head, or broad part, is directed anteriorly. It is continuous with the anterior perforated space. The tail terminates in the nucleus amygdalae. The
caudate nucleus and the lenticular nucleus are connected by bands of fibers
both at their anterior and their posterior extremities. The presence
of considerable numbers of fibers, which pass from the caudate nucleus
to the lenticular nucleus, together with the position of the nucleus in
the floor of the ventricle, renders probable the view that the caudate
nucleus is derived from the epistratum in fishes. The caudate nucleus
still receives many olfactory impulses, as is to be expected when one remembers
that its primitive function is concerned in olfactory coordinations.
The presence of the optic radiation and the cortical structures is associated
with increasing complexity of relations among the nuclei of the striate
and thalamic bodies.
The nucleus lentiformis, or the lenticular nucleus, is composed of a lens-shaped mass of gray matter, crossed by many sheets of thin white matter. It is thus divided into three zones, of which the outer is called the putamen. The two inner masses are rather less well supplied with blood, contain less pigment, and are thus less pale than the other gray masses. They are called the globus pallidus for this reason. The putamen is characterized by the great size of its multipolar cells and by the great length and irregularity of their dendrites. Its gray matter is continuous with that of the caudate nucleus anteriorly. It both gives and receives fibers from the caudate nucleus and from the globus pallidus. The globus pallidus is composed of two, three or more masses of gray matter. In this part of the brain the circulation is less plentiful than in other parts of the gray matter. The cells of the globus pallidus are smaller and less irregular in outline than in the putamen. The globus pallidus is less well developed in the lower vertebrates than are the other masses composing the corpora striata. The connections of the different parts of the corpora striata are either very much alike or they have not been well worked out; for this reason it seems best at present to give them in one group. Each corpus striatum receives fibers from the following sources: I. Olfactory axons, of the second or higher orders, terminate in the caudate nucleus and the putamen. II. Fibers from the thalamus, midbrain, and the sensory nuclei of the second and higher orders, enter the lenticular nucleus probably in all of its parts, to a certain extent. III. Descending fibers from all parts of the cortex of the same hemisphere enter the striatum. IV. Collaterals from the descending cortical fibers enter the putamen, and probably the globosus and the caudate nucleus. This is most conspicuously seen in descending pyramidal fibers from the precentral convolution. V. Fibers from the thalamus and the striatum of the opposite side enter the striatum. VI. Fibers from the red nucleus probably terminate in the inner section of the pallidus. Such fibers would make a part of the indirect sensory pathway. The fibers leave the striatum to pass to the following destinations: I. Fibers pass in the strio-thalamic bundles to the thalamus of the same side. II. Fibers pass to the thalamus of the opposite side. III. Fibers pass to the hypothalamic centers and to the red nucleus and substantia nigra of the opposite side, and also to the same side. IV. Fibers pass upward to the cerebral cortex of all areas. The stimulation of the various areas of the corpora striata gives no recognizable results. Diseases
of the striatum are almost certain to affect the internal capsule, and
to produce the paralysis due to this injury. Injury of the striatum
without injury to the capsule may produce no symptoms, or may give rise
to indefinite and variable symptoms, so that no conclusions can be drawn
therefrom, nor can the diagnosis of the lesion of the striate body alone
be made antemortem.
This nucleus is produced by a thickening of the gray matter of the cortical infolding associated with the hippocampus major. It thus protrudes upon the surface of the inferior horn of the lateral ventricle as an almond-shaped swelling. It is of interest in its position in phylogeny as one of the very earliest cortical developments, and it is in all vertebrates one of the chief cortical centers of the sense of smell. The end of the tail of the caudate nucleus is continuous with its gray matter. It receives fibers from the olfactory cells of the second and higher orders, and perhaps from neighboring areas of the cortex. A few fibers of long association tracts bring this area into relationship with the other cortical areas in mammals. It sends the axons of its cells to the following destinations: I. To the nucleus habenulae by the stria medullaris, or tractus olfacto-habenularis; II. To the hypothalamus by way of the taenia semicircularis, or lateral olfacto-hypothalamic tract; III. To the hippocampus major by way of the subcallosal gyrus and the fimbriae; IV. To other areas of the cortex by way of the long and the short association tracts; V. To the opposite side of the brain by way of the several commissures, either directly or indirectly. |
{kind=link}
{kind=link}